2013年11月28日


左から上村教授、服部特定助教
服部佑佳子 生命科学研究科特定助教、上村匡 同教授らの研究グループは、神経細胞の多様性を生み出すしくみについて解明しました。この研究成果は、2013年11月28日に米国科学誌「Developmental Cell」誌のオンライン版に掲載されました。
概要
神経細胞(ニューロン)と一口に言っても、極めて多様な個性を持つ種類(サブタイプ)が存在します。神経系の働きにより個体の正常な活動が支えられるのは、サブタイプごとに選択的な生理機能を分担して果たしているからです。そしてそれぞれの生理機能を果たすには、サブタイプごとに特徴的な形状の神経突起を発達させることが必要です。研究グループは、形態的に異なる特徴を生み出す二つの転写調節因子の働きを研究しました。そして、それらの転写調節因子が支配する遺伝子発現プログラムを、全ゲノムにわたって解明し、そして互いに比較することで、サブタイプ間の形態的なちがいを生み出す原理を発見しました。
背景
多細胞生物の初期発生では、様々な細胞が正確に生み出され、器官や体を作り上げていきます。器官の中でも、特に神経系は多様なニューロンのサブタイプによって構成され、サブタイプごとに機能を分担する分業体制が高度に発達しています。ニューロンのこの多様性はどのようにして生まれるのでしょうか。これまでに、同じ神経幹細胞から、発生の時期によって異なるグループの子孫細胞が生み出されるしくみなどの理解は進んでいました。一方で、誕生直後のニューロンはまだ十分に個性を獲得して(分化して)おらず、神経系全体として機能の細分化を達成するには、ニューロンが生まれた後に働くしくみも必要です。生まれたニューロンがどの転写調節因子を発現するかで、異なるサブタイプに分化する観察事実が、我々を含め、世界中のグループから報告されてきました。しかし、それらの転写調節因子が、実際にどのような遺伝子の発現を調節してサブタイプごとの個性が獲得されるのか、そして異なる転写調節因子の間で、支配する遺伝子発現プログラムがどの程度違っているのか(裏返せばどれだけ共通なのか)は、断片的にしか分かっていませんでした。
研究手法・成果
手法(用いたモデル系とゲノムワイドな手法)
神経細胞においてシナプスや感覚入力の処理を支える樹状突起の形態は、サブタイプごとに非常に多様であり、生理機能の違いに大きく寄与しています。ショウジョウバエ感覚神経のひとつdendritic arborization(da)ニューロンは、表皮と筋肉の間に2次元的に樹状突起を展開しており、樹状突起のサイズと複雑度により四つのクラスに分類されます。クラス I ニューロンは最も小さく単純なくし形の樹状突起を持ち、対照的にクラス IV ニューロンは最も大きく複雑な、放射状の樹状突起を発達させます(図1)。クラス I ニューロンにおいて、くしの歯に相当する突起は身体の前後軸に沿って伸びており、ショウジョウバエの幼虫はこの突起を介して身体の伸展運動を感知しています。一方、幼虫は、皮膚の裏側を隙間なく埋めるクラス IV の突起を介して、針で刺された場合や熱などの痛みを伴う刺激を感知し、忌避行動を示します。研究グループはこれまでに、転写調節因子Abrupt(Ab)がクラス I で、Knot/Collier(Kn)がクラス IV で選択的かつニューロンの誕生後に発現することで、それぞれの樹状突起形態の発達に貢献することを明らかにしました。このようなサブタイプの特徴付けをする転写調節因子を、サブタイプセレクターと呼びます(図1)。
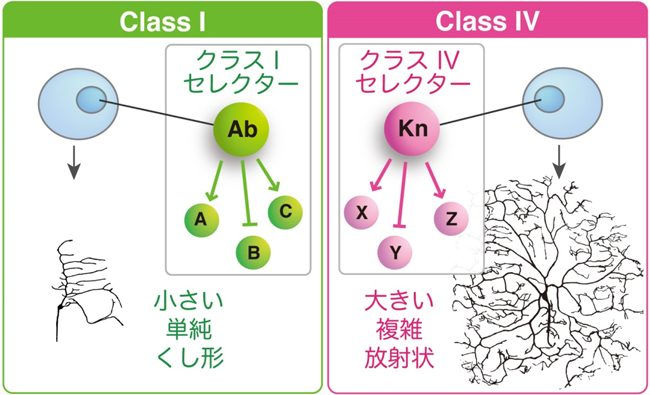
図1: クラスIおよびクラス IV ニューロンの樹状突起形態とサブタイプセレクター。転写調節因子Abruptはクラス I で、Knotはクラス IV で選択的に、かつニューロンが誕生した後になって発現され始め、それぞれのクラスに特徴的な樹状突起の形成に機能している。これらの転写調節因子の標的遺伝子や、標的遺伝子による突起形成メカニズムはほとんど分かっていなかった。
本研究では、二つの転写調節因子を2種類の全ゲノムにわたる(ゲノムワイドな)手法により解析することで、転写調節プログラムを明らかにしました(図2)。まず、AbやKnのゲノム中での結合部位を網羅的に探索し、結合する遺伝子群を同定しました。またこの手法と並行して、磁性ビーズを用いてdaニューロンを単離し、遺伝子発現プロファイルを調べることで、AbやKnによって発現制御を受ける遺伝子を同定しました。これらの結果は、実験データを統計的手法で解析することで得られました。
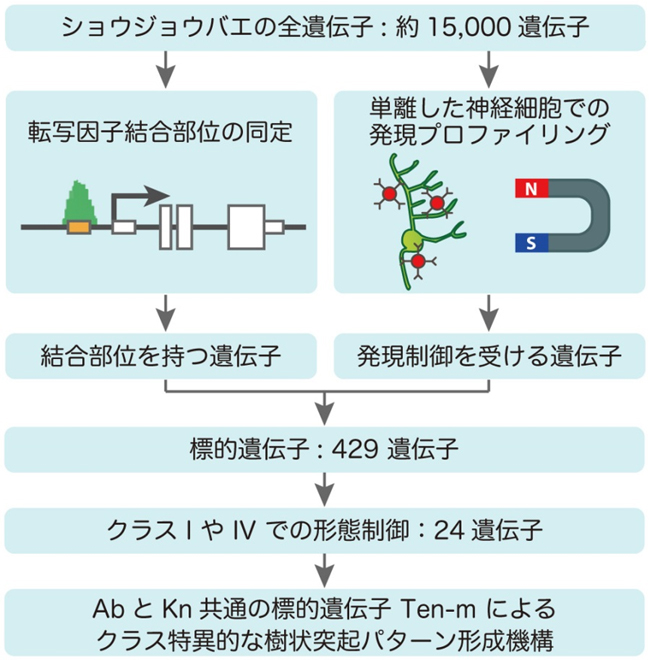
図2: AbruptとKnotの転写調節プログラムをゲノムワイドに解析した研究の流れ。全ゲノムにわたって、AbruptとKnotの結合部位や標的遺伝子を同定し、互いを比較し、AbruptとKnotの転写制御において何が異なることで、クラス間の形態的なちがいが生み出されるのかを明らかにした。
成果
上記の結果を統合し、ショウジョウバエのゲノム中に存在する約15,000遺伝子の中から、AbとKnが結合し、かつ発現制御を行う遺伝子(標的遺伝子)を合計429個同定しました(図2)。この中で、56遺伝子がAbとKnに共通でした。AbとKnの結合プロファイルを調べたところ、どちらも神経発生への関与が示唆される遺伝子の、細胞タイプ特異的エンハンサーに濃縮していることが分かりました。
次に、同定した標的遺伝子のクラス I やクラス IV ニューロンでの発現を阻害すると、樹状突起の形態に顕著な異常を生じる24の遺伝子を同定しました。これらの遺伝子がコードしているタンパク質の分子機能は、カルシウムイオンチャンネル、膜輸送制御タンパク質、mRNA結合タンパク質など多岐にわたっており、このような様々な分子機能をもつ遺伝子群が協調してはたらくことで、それぞれのクラスの樹状突起形態が生み出されていることが分かりました(図3)。
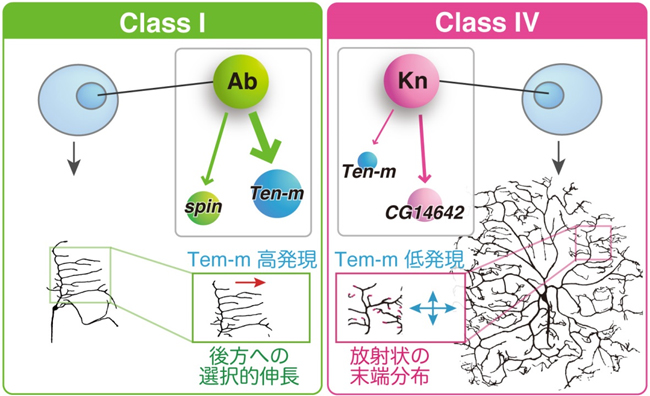
図3:クラス I とクラス IV ニューロンの樹状突起形態に機能する標的遺伝子群の同定。たとえば、Abrupt特異的な標的遺伝子であるspinはクラス I ニューロンで、Knot特異的な標的遺伝子CG14642はクラス IV ニューロンで発現し、それぞれのクラスでの樹状突起形成に機能していた。一方、AbruptとKnotに共通の標的遺伝子Ten-mは、両方のクラスで発現して突起形成に機能していた。ただし、共通する標的遺伝子の中には、Ten-mのようにクラス間で発現量に差がある遺伝子が複数見つかった。Ten-mの場合、この発現レベルの差がクラス間での突起形態のちがいに重要であることがわかった。
さらに研究グループは、AbとKnの標的遺伝子がどのようにしてクラス特異的な樹状突起形態を生み出すのか、その分子メカニズムを明らかにすることを目指しました。そのために、AbとKnに共通の標的遺伝子であるTen-mに着目して解析しました(図4)。Ten-mは哺乳類まで保存されたTeneurinファミリーに属する同種親和性の細胞接着因子です。発現パターンの解析から、クラス I ニューロンでは強く、クラス IV ニューロンでは弱く発現していることがわかりました。また興味深いことに、樹状突起と接触する表皮細胞で、各体節の中央(ニューロンの細胞体が位置する)から、体節境界付近(ニューロンの突起末端が位置する)にかけて、Ten-m発現レベルが変化していることがわかりました。
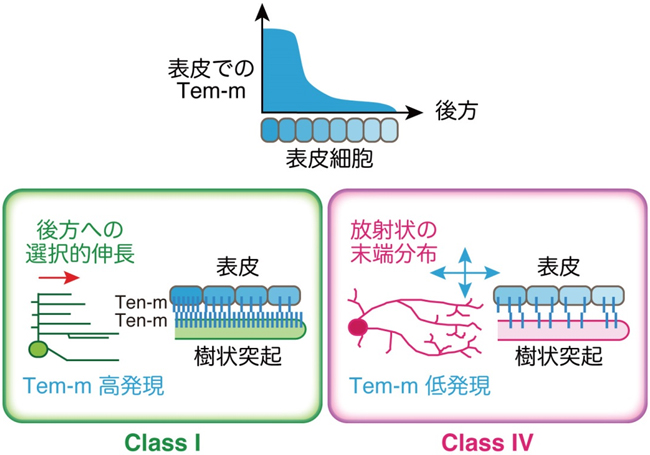
図4:Ten-mによるクラス特異的樹状突起形成メカニズム。Ten-mの発現レベルはクラス I ニューロンでは高く、クラス IV ニューロンでは低い。このクラス間の発現レベルの差が、周辺組織である表皮での空間的な発現レベルの強弱と協調して、それぞれのクラス特異的な突起パターン形成に機能している。
そこで研究グループは、表皮と樹状突起との相互作用が、樹状突起形態形成に寄与しているのではないかと考え、以下のように検証を行いました。表皮でのTen-mの発現を阻害したり、表皮でのTen-mの空間的な強弱パターンを乱したりすると、クラス I やクラス IV の突起形態に顕著な異常を生じました。さらに、クラス間でのTen-mの発現レベルのちがい(クラス I >>クラスIV)が、クラス間での突起パターンのちがいに寄与しているかを調べるために、通常Ten-mの発現量が低いクラス IV ニューロンで、Ten-mを強制的に過剰に発現させました。その結果、通常、放射状のいろいろな向きに分布するクラス IV の突起末端が、クラス I のように体の後方へ選択的に枝を伸長させる表現型を得ました。これらの結果から、クラス間でのTen-mの発現レベルの強弱が、周辺組織のTen-m発現レベルの空間的な強弱と協調してはたらくことで、それぞれのクラスに特徴的な突起パターンが形成されていることが明らかになりました(図4)。
さらに研究グループは、AbとKnに共通の標的遺伝子の中に、Ten-m遺伝子以外にもクラス I とクラス IV の間で発現レベルに顕著な差のある遺伝子を同定しています。これらの遺伝子も、それぞれのクラスに特徴的な突起形状の発達に寄与していることが想像できます。以上の結果から、神経系が発達する際には、それぞれのサブタイプの分化に特異的な遺伝子と、共通に用いられる遺伝子がどのように使い分けられて、サブタイプ選択的な形態的特徴を生み出しているのかを明らかにしました。
波及効果
本研究により、二つのサブクラスセレクターの標的遺伝子のゲノムワイドなリストが公開されました。サブクラスセレクターと標的遺伝子の組み合わせの中には、種を越えて保存する例も知られており、本研究の成果はヒトの複雑な神経系を構成する特定のサブクラスの誕生の解明につながります。さらには、他のサブクラスセレクターについても本研究と同様の解析を進めることで、任意のサブクラスの誕生をプログラムしたり、特定のサブクラスの欠損によって生じる障害を他のサブクラスで補うことも実現するかもしれません。また、神経系のみならず他の器官における細胞分化過程も、本研究と同様のアプローチが可能であるため、生命科学の多方面の研究に貢献できます。
本研究成果は、文部科学省科学研究費補助金・新学術領域研究「メゾスコピック神経回路から探る脳の情報処理基盤」および日本学術振興会・特別研究員制度の支援によって得られました。
用語解説
転写調節因子と標的遺伝子
ゲノムDNAの特定の箇所に結合して、周辺の遺伝子の発現を高めたり、または抑制したりするタンパク質。発現調節を受ける側の遺伝子を、標的遺伝子と呼ぶ。
樹状突起
神経細胞は2種類の突起構造を発達させる。シナプスを介する入力や感覚入力(痛み、臭い、音など)を受容するのが樹状突起であり、神経細胞のサブタイプごとにその形状(突起全体の大きさや分岐の複雑度など)は様々である。
細胞接着因子
細胞同士、または細胞と細胞外基質との接着を担うタンパク質。細胞膜を貫通する(細胞膜に突き刺さっている)タンパク質であることが多く、細胞外領域で互いに結合し、細胞同士を接着させる役割を持つのが同種親和性の接着因子である。
書誌情報
[DOI] http://dx.doi.org/10.1016/j.devcel.2013.10.024
[KURENAIアクセスURL] http://hdl.handle.net/2433/179534
Yukako Hattori, Tadao Usui, Daisuke Satoh, Sanefumi Moriyama, Kohei Shimono, Takehiko Itoh, Katsuhiko Shirahige, Tadashi Uemura
"Sensory-Neuron Subtype-Specific Transcriptional Programs Controlling Dendrite Morphogenesis: Genome-wide Analysis of Abrupt and Knot/Collier"
Developmental Cell 27, pp1-15
Available online 27 November 2013
- 京都新聞(11月28日 25面)および日刊工業新聞(11月28日 25面)に掲載されました。